Single Cell
原理参考 10x Genomics Support
oligo文库注:Cell Hashing 技术基于 CITE-seq,可同时测定单细胞RNA、表面蛋白标记物,情形类似:
## 每个单克隆抗体都与一个短DNA寡核苷酸结合,被称为hashtagoligo(HTO)==AntibodyBarcode
CapturedRNA--------UMI-----------CellBarcode---
|--Beads
CapturedProtein-----Antibody-----HTO-----------
scRNA
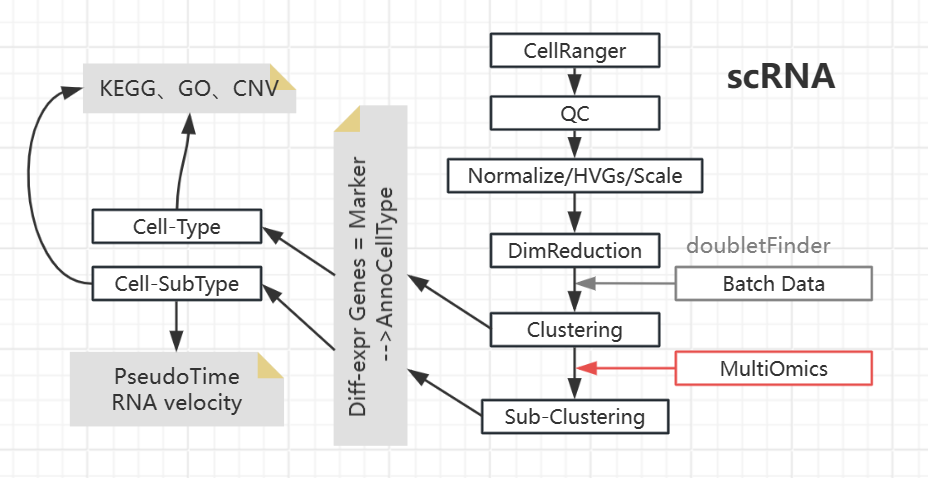
| 任务 | 工具 |
|---|---|
filtered_feature_bc_matrix/ |
CellRanger |
| PIPELINE | R: Seurat Py: Scanpy |
| CellType Anno | R: SingleR, Metacell, Cellassign |
| Batch Integration | R: Seurat, Harmony, LIGER |
| MultiOmics Integration | R: Seurat |
| Doublet Detection | R: DoubletFinder, DoubletDecon Py: DoubletDetection, Scrublet |
| PseudoTime(Traj) | R: monocle3 Py: Scanpy(paga) |
| RNA Velocity | Py: scVelo |
| GO GSEA / KEGG Enrichment | R: clusterProfiler |
| SeuratObj <--> Scanpy_h5ad | Seurat: Convert() |
此外,inferCNV 分析CNV以判断是否是肿瘤细胞,hdWGCNA 分析基因模块,SNV分析(META-CS实验技术改进),...
Bulk mRNA 数据可以帮助验证scRNA数据的完整性;如果还有其它bulk多组学数据,也可以尝试算一下相关性
R:Seurat+SingleR
- Seurat对每个细胞统计符合pattern的基因量,然后可以根据这个统计量进行Filtering,或者在Scale时候消除这个统计量的影响(RegressOut),e.g.
pbmc[["percent.mt"]] <- PercentageFeatureSet(pbmc, pattern = "^MT-") ## Calculating
pbmc <- subset(pbmc, subset = nFeature_RNA > 200 & nFeature_RNA < 2500 & percent.mt < 5) ## Filtering
pbmc <- ScaleData(pbmc, features = rownames(pbmc), vars.to.regress = "percent.mt") ## RegressOut
| Data QC | 说明 | pattern |
|---|---|---|
| Filter off Genes | Appear in >3 Cells | -- |
| Filter off Cells | Not Doublet: via Detectors minGeneNum > 200-500 maxGeneNum < X, X decided via 'VlnPlot()' (Doublet may contain more genes) |
-- |
| Filter off Cells + RegressOut | Mitochondrial Genes <15% | "^MT-" |
| Filter off Cells + RegressOut | Ribosomal Genes <5% | "^RP[SL]" |
| Filter off Cells + RegressOut | Hemoglobin Genes "HBA1","HBA2","HBB","HBD","HBE1","HBG1","HBG2","HBM","HBQ1","HBZ" |
^HB[^(P)] |
| Filter off Cells + RegressOut | Cell-Cycle: 见下文 | cc.genes + CellCycleScoring() |
-
根据Variance选出HVGs: GeneA_cellA v.s. GeneA_Overall
- Opt. If dataset is large, use HVGs for afterward analysis
-
(Opt.)根据HVGs进行 Cell-Cycle Scoring and RegressOut:Pick HVGs --> Load privided Cell Circle Genes
cc.genes--> CellCycleScoring() --> ScaleData()- Opt.1: vars.to.regress = c("S.Score", "G2M.Score")
- Opt.2: vars.to.regress = c("CC.Difference");CC.Difference = S.Score - G2M.Score
- 注意,Opt.1不适合关注细胞分化的实验,因为会完全消除非周期细胞和周期细胞之间的区别;Opt.2 将保持二者间细胞周期差异,仅消除二者内部细胞周期差异
- Tips:可以通过查看对PC贡献最多的基因是否是细胞周期相关基因来判断影响
-
随后进行Clustering,假设每个Cluster代表一种Cell-type;再次对Cell-type进行Clustering,可得到细胞的分型。
- Find marker genes --- Diff-expr genes: cluster v.s. clusters
- Annotate clusters based on marker genes
| -- | DB for SingleR() | -- | Tips |
|---|---|---|---|
| Cell-Type | HumanPrimaryCellAtlasData(label.main), CellMarker, PanglaoDB, SingleR datasets, Papers | e.g.Tcell/Bcell/... | 在这个水平上进行一些常规的分析(cluster间的比较),比方说KEGG、GO富集分析,差异基因表达分析(cell-type间) |
| Cell-Subtype | MonacoImmuneData(label.fine) | e.g.对CD4细胞细致分类 | 在这个水平上进行一些更加细致的操作,比如:拟时序分析,RNA velocity |
Meta
对宏基因组使用scRNA与Basic Pipeline不同;Basic Pipeline中已知样本来自某一生物,所以可以很简单的使用参考基因组进行每个细胞内、每个基因的定量;而宏基因组的潜在参考基因集过于庞大,耗费过量资源;且宏基因组使用scRNA技术的目的之一是希望得到更准确的细胞组成信息(现有Binning有时准确度堪忧),需求是对每一个cell进行准确的物种注释。
为了尽量缩小潜在参考基因集范围,可以参考相似研究的Meta Bulk注释提到的基因组;Tips: GTDB使用ANI进行分类,可以使用本地储存的文件,不用去NCBI再下载
其单细胞QC步骤一如前文所述。有两种或许可行的注释、定量方法:
- Anno via FastANI:当需要注释的细胞数量较少时,可以将其比对参考DB,根据ANI确定可能Taxa范围
- Anno via Mapping:通过cdhit等对参考基因集进行去重,然后将scRNA数据map上去,根据细胞的mapping信息决定其物种注释(也许可以使用cellranger)
Tips:有时如果物种注释结果不理想,我们也可以进行单细胞水平的功能注释(GO,ARG,pathway...)
STOmics
BGI STOmics 的v1实验流程与 10X Genomics 类似,将磁珠(beads: CellBarcode_UMIs)与细胞裹入同一液滴中,液滴回收、破乳、反转录、扩增、测序,所得到的reads可以带有beads上的信息。理想状态下 1液滴=1磁珠+1细胞,但如果bead浓度太低则部分液滴中无bead,导致细胞回收率低;但如果bead浓度太高则1液滴中有多个beads,在后续分析中会被标识为多个细胞。(相应的,如果细胞浓度过高,则1液滴中有多个细胞,即Doublet)
BGI STOmics 的v2实验流程改进为使用2种磁珠,大磁珠(CellBarcode_UMIs)可捕获RNA/oligo、同时可结合环境中的各种小磁珠(Index_UMIs)。1液滴=n小磁珠+n大磁珠+1细胞,位于同一液滴的大磁珠具有相似的小磁珠结合pattern(因空间靠近、微环境相似),于是可基于此信息将大磁珠捕获的RNA合并回完整的Cell;如是通过增加磁珠浓度提高细胞回收率。
情形类似:
|---SmallBead_IndexA
CapturedRNA--------UMI-----------CellBarcode--- |---SmallBead_IndexA
|-- BigBead --|---SmallBead_IndexC
CapturedProtein-----Antibody-----HTO----------- |---SmallBead_IndexB
|---SmallBead_IndexA
rawReads数据处理的流程:QC/UMI Filtering-->Mapping-->PISA Gene anno-->Beads Filtering-->Beads Merging-->Gene_Expr_Mtxs,格式同CellRanger的结果
scATAC
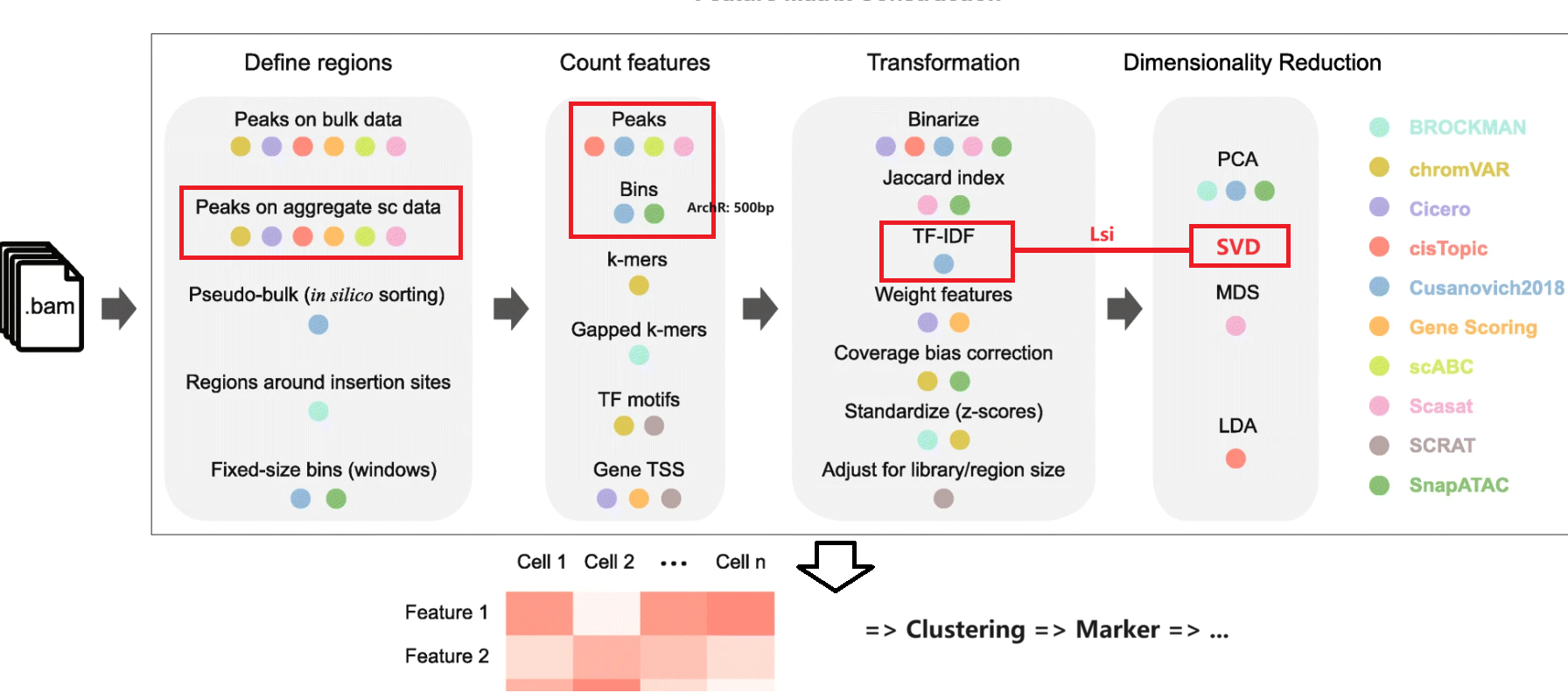
推荐ArchR(Bin)一站式分析。此外常见Signac(Peak)+Seurat。
Feature Matrix
已知reference情况下,可通过 CellRanger 得到每个细胞的 Peak 信息。由于Peak数据一般十分稀疏,因此某些处理流程会选择合并,例如 Ansuman T. Satpathy (2019) 的QC流程:
1. 去除低质量细胞
- TSS enrichment score 指所有基因TSS位点测序深度的平均值,一般取TSS上下游1000-2000bp的值为基准
- Unique fragment > 1000
2. 降低分辨率:2.5kb为一个bin合并Peak数目,生成TileMatrix
3. 生成 Pseudo-bulk ATAC 数据:根据TileMatrix的聚类信息合并而成
4. 对每一个 Cluster 所对应的 Pseudo-bulk 数据进行 Peak Calling (MACS2)
5. 根据bulk得到的Peak信息,选取单细胞数据中的高质量Peaks
- scATAC一般不直接Normalize,而是使用TF-IDF挖掘Feature
- Doublet
- ArchR: 模拟双细胞,随后在低维空间中识别与之相近者为双细胞
- scDblFinder
- Batch Integration: Harmony, LIGER
CellType Anno
Clustering后,10x 提供关于CellType Anno的建议:
Opt 1. 寻找Cluster间有accessibility差异的Peaks,其下游最近的基因为Marker Gene
Opt 2. 用户自定义 Cell Type-Specific Feature Set。常见将Peak Matrix 转换为 Gene Scores Matrix(可以理解为基于表观数据预测出的基因表达值), 随后用SingleR 等进行细胞类型注释
Opt 3. Seurat TransferData 将携带注释的scRNA数据与scATAC数据在低维空间对齐
Downstream
得到Marker Peaks/Genes 后:
| Task | 说明 | Tool |
|---|---|---|
| ... | GO/KEGG、轨迹分析、细胞速率、... | - 同scRNA - GREAT有些类似对差异Peak进行GO Term分析 |
| 调控网络分析 | Peak-Peak Co-accessibility | - Cicero - Pando+scRNA - ArchR |
| Motif Analysis | 寻找某个TF的 binidng site 的特征基序(随后可以在全基因组范围内寻找符合motif的潜在binding site) | chromVAR |
| Footprint Analysis | 潜在的TF binidng site是否结合TF:如果某一开放区域已经结合TF,则此区域不会被测得,表现为peak中间有凹陷 | TOBIAS |
参考
各种单细胞测序技术: https://zhuanlan.zhihu.com/p/569346733
数据集汇总: https://www.jianshu.com/p/49e5cf91f819
NBIS: https://www.jianshu.com/p/1e29e3b9a4ab
Doublet: https://zhuanlan.zhihu.com/p/376439628
HVGs: https://www.jianshu.com/p/3d40c56e5fc8
Cell Circle: https://zhuanlan.zhihu.com/p/82654538
Batch: https://www.jianshu.com/p/ebc328f9fb73
GREAT: https://www.jianshu.com/p/7f6ce4f238cb